The following account began in a 1998 IPA field guide to East Java, written by Peter Lunt. Parts of it were copied, un-cited, in a 2002 paper for which an apology was received. It was updated in Lunt and Allan (2004) and Lunt (2013a). If you need to cite any of the material below I recommend Lunt (2013a).
Introduction
Understanding the Letter Stages is important to comprehend work on the whole of eastern Tethys. While many have pointed out that they are not true Stages, like the Chattian or Aquitanian, but biozones, very few have advocated replacing these popular and orderly-named zones with cumbersome fossil-zonal names. For the most part the application of these Letter Stages has been consistent over the last fifty years (with the exception of the Upper T.f). Also, the Letter Stages appear to represent assemblages between faunal turnovers, so it makes sense to conserve them as practical stratigraphic divisions. They should be considered as originally intended, as being of a similar scale and replacements for the “Eocene”, “Miocene” and “Pliocene’ etc., Epochs, that were also first defined on fossil content but are now used as names for chronostratigraphic divisions.
The Letter Stages were originally defined on associations of certain microfossils; that is, the contents of the units are defined. This contrasts with planktonic microfossil zones that are defined by their boundaries, which are single species evolution or extinction datums. Ideally planktonic zonal datums should correlate well and approximate time-lines. However in larger foraminiferal carbonate sequences there is a chance that Letter Stage boundaries are delimited by facies changes, therefore correlation lines or the concept of “datum planes” are considered inappropriate for the larger foram biozones.
The Epochs of life
The term Tertiary first appeared in the eighteenth century, in a four-fold stratigraphic subdivision comprising Primary, Secondary, Tertiary, and Quaternary units (old Latin words for First Second, Third and Fourth). The first referred to crystalline rocks and the second to hard, stratified rocks with some fossil content. The third, Tertiary, unit was recognised as hardly indurated sediments that were obviously not part of young soils or alluvium. The similar term Cainozoic (or Cenozoic; from the Greek kainos = new and zoon = life) was defined in the mid nineteenth Century on broad biostratigraphic principles, being the sediments (and their fossils) deposited in the period after the end-Cretaceous mass extinction. It includes both the Tertiary and Quaternary.
Just prior to the definition of the Cainozoic, Lyell (1833) divided the Tertiary of Europe into the Eocene, Miocene, Older Pliocene and Newer Pliocene (the latter pair revised in 1839 into Pliocene and Pleistocene). As with the Cainozoic these were biostratigraphy-based units, as reflected in the Greek roots of the names, viz.: Eocene = “dawn of the new”, Miocene = “less new”, Pliocene = “more new”, Pleistocene = “most new”. Lyell defined his Eocene as strata with less than 5% of mollusc fossils still living, the Miocene as strata with about 17% extant forms, and the Pliocene as strata with 35-50% extant forms. (The Paleocene* and Oligocene were added in 1874 and 1854 respectively as workers discovered significant sections of sediments at the base of the Eocene and between Lyell’s Eocene and Miocene.)
It is worth noting that at about the time Lyell defined these ages, W.B. Carpenter had just published his microscope studies of Nummulites and other genera (1850) and clarified the structure of these fossils, although they were then placed in the Cephalopoda and thereby considered a form of mollusc (but not used in Lyell’s percentage method). By the end of the 19th Century the first synthesis of European stratigraphy, at Epoch or Series level, was virtually complete and work was then focused on applying these Epochs in the relative dating and correlation of the lithostratigraphic stages defined on various stratotypes across Europe.
One of the many over-lapping terms proposed for the older Tertiary was the Nummulitique of Emile Haug, proposed in 1902 for the Palaeogene (Palaeogene itself introduced in 1853 but only becoming a practical term in 1874 when it could cover a combined Paleocene and Eocene). The term Nummulitique survived for some fifty years and is directly comparable to the more diversely fossiliferous Indonesian Tertiary a through d. This wider use of the term Nummulitic was one of the reasons that the name Nummulites was adopted in preference to the original (senior) term Camerina.
Subdivisions of the Tertiary in SE Asia
In Indonesia, where the Tertiary is a much more significant part of the sedimentary geological record than it is in Europe, the 19th and early 20th Century workers attempted to utilise the new European stratigraphic schemes. Professor K. Martin (1852-1944), studying the mollusc faunas of Java and surrounding areas was particularly outstanding in this field. His work was able to follow that of Lyell in Europe in utilising the ratios of extant to extinct faunas to indicate relative ages. Two major contributions to Tertiary stratigraphy came from the work of Martin. Firstly he argued that the mollusc faunas of the East Indies developed separately from those of Europe and secondly, as a consequence of this, the stage names of Europe could not be correlated with confidence to the Indonesian region.
The birth of the Letter Stages
Martin’s view that the Indonesian area could not be correlated with the European stratigraphy gradually won acceptance between the first publication of Martin on the famous Eocene section at Nanggulan in 1914, and the comprehensive review of Indonesian geology by Rutten in 1927. This break with European stratigraphy was the reason the Geological Survey of the Dutch Indies began to set up their own stratigraphic system, and there is fair reason to acknowledge the Nanggulan section as the place of origin of the Letter Stages. There is, however, a twist to this tale that is not widely known.
The original Nanggulan samples were collected by a coal prospector, van Dijk in about 1872, but these were a mixed bag from the Eocene inlier and a Miocene formation (Jonggrangan Fm.) that unconformably overlies the older beds. Consequently the first work by Martin, used by Verbeek and Fennema (1896) in their review of Java and Madura, arrived at a value of 28% extant species. This was considered a typical Miocene ratio, but with the old Tertiary foraminifera Discocyclina and Nummulites present, a compromise age of Oligocene age was assigned to the Nanggulan beds. In 1920 Martin did his own field work and began a number of detailed descriptions of the exceptionally well preserved fauna of the site (106 gastropods, 23 bivalves, 3 scaphopods and 4 foraminifera, – the most diverse old Tertiary mollusc fauna in the east Asian region). However no species at Nanggulan were considered extant, in contrast to the 5% of the Eocene molluscs in Europe, plus it contained some genera not known from Europe before the Oligocene. Martin asked the expert on the Eocene of the Paris Basin, Cossman, to compare the faunas (cf. Rutten 1927, chapter VI and Oppenoorth & Gerth 1929). Cossman confirmed Martin’s observations that the Nanggulan fauna consists of forms closely related but not identical with those of the Paris Eocene. This observation “led Martin to the conclusion that the fauna of Nanggulan belongs to the younger Eocene, but that a common connection between the European part of the Tethys geosyncline and the region now occupied by the Indian Archipelago no longer existed.” (p.5 Oppenoorth & Gerth).
The evolution of the Letter Stages
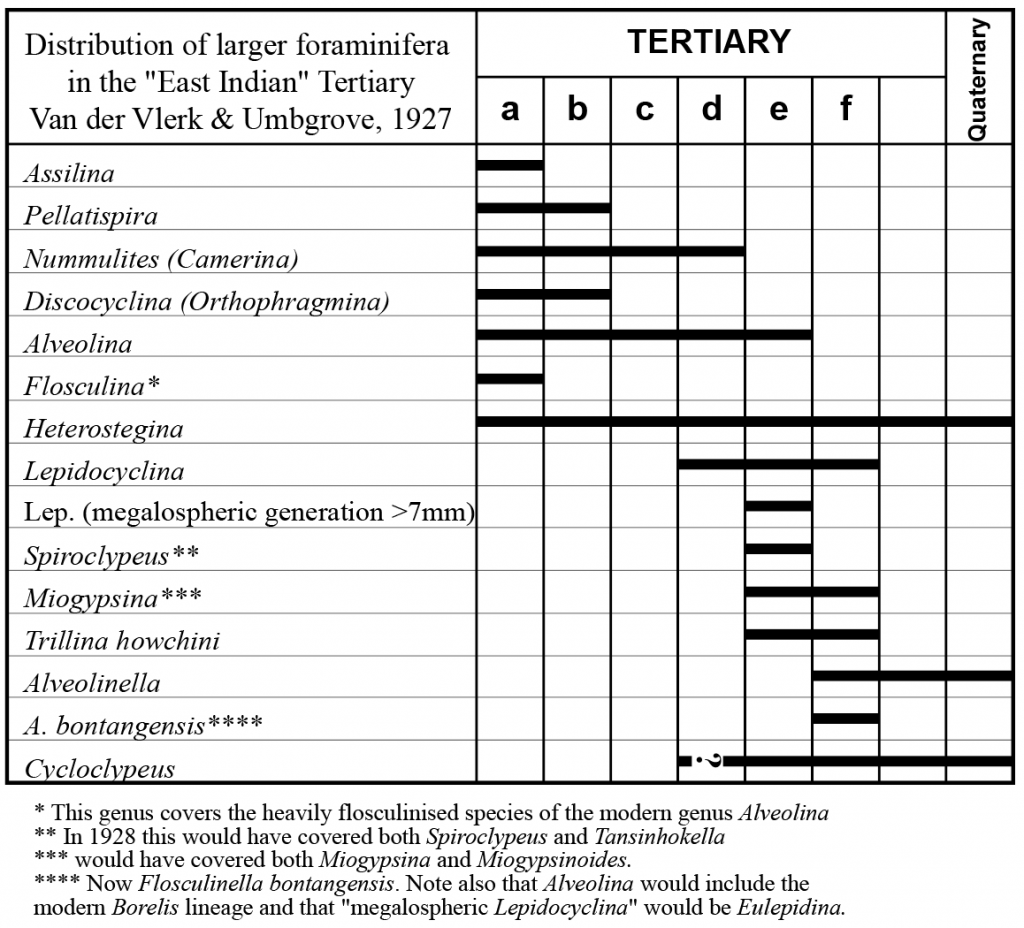
In 1927 Van der Vlerk and Umbgrove published the Letter Classification of the Indonesian Tertiary, based on larger foraminifera. This scheme subdivided the Tertiary into seven parts. Six parts were labeled “a” through “f” (e.g. Tertiary “a”, or “T.a” for short), and a seventh section of non-orbitoidal Tertiary between Tf and the Quaternary was noted. This scheme was immediately successful and was rapidly adopted as a workable biostratigraphic scheme for the the Far East, even if the correlation with the European epochs and stages was not adequately known. The authors pointed out that the taxonomy and detailed biostratigraphy of the genera used was not fully understood, and suggested that future work would increase the number of recognisable subdivisions.
It is ironic that this break from European stratigraphy may not have been necessary. Reviews of the Eocene faunal ratios first described by Lyell, summarised by Morley Davies (1935, revised by Eames, 1975) arrived at the conclusion that “even in the Oligocene [separated from the top Eocene after the time of Lyell’s first work], Recent species never exceed 1 per cent” of fossil forms. While mollusc studies had been important in understanding the stratigraphy of the Indonesian area, and specialists like Martin could reliably arrange faunas in stratigraphic order based on ratios of extant species, it was the larger foraminifera that were to be the foundation of the new stratigraphic system. By the late 1920s they had become the preferred fossil group for biostratigraphy, primarily as they were more abundant than molluscs, but also they did not require extensive knowledge of large numbers of living and fossil species. A scheme was developed that utilised assemblage zones rather than percentages of extant forms. The larger foraminifera assemblage zones could be identified by the presence of a few key taxa, often with a hand-lens in the field.
The original proposal of the Letter Stages was:
Tertiary a, the oldest larger foraminifera assemblage known in the region, characterised by Nummulites, Discocyclina, Pellatispira, “Alveolina” (i.e. the forms that now fit under Alveolina s.s. [or Fasciolites of some authors]) “Flosculina” (i.e. what would now be regarded as heavily flosculinised species of Alveolina), and Assilina.
Tertiary b faunas also contain Nummulites, Discocyclina, Pellatispira and “Alveolina” (which in this case would be the plexus that later became known as Clausulus, then Neoalveolina’, but now under the senior name, Borelis). This stage differs from the Tertiary a by the absence of Assilina and “Flosculina”.
Tertiary c contains Nummulites but Discocyclina, and Pellatispira are absent.
Tertiary d still contains Nummulites, overlapping with the first appearance of the lepidocyclinid forms. Van der Vlerk and Umbgrove divided their lepidocyclinid forms into two groups. “Lepidocyclina” and “Lepidocyclina megalospheric generation, tests >7mm)”. These taxonomic concepts equate to Lepidocyclina(isolepidine, nephrolepine and related forms) and Eulepidina respectively. These “megalospheric generation >7 mm” forms are noted to have a limited stratigraphic range that is identical to Spiroclypeus. It is surprising that the term Eulepidina was not used as it had already been proposed in 1911 by Douvillé working in the Philippines.
Tertiary e is identified by the “Lepidocyclina megalospheric generation” (=Eulepidina) and Spiroclypeus mentioned above, as well as by Miogypsina, a term that in 1927 included both the modern Miogypsina and Miogypsinoides, and by “Trillina howchini” (= the whole genus Austrotrillina at that time).
Tertiary f is characterised by Alveolinella – which then included “A.” bontangensis (= the modern Flosculinella bontangensis). The forms Eulepidina andSpiroclypeus were absent from this stage as was “Alveolina” (Borelis forms).
Shortly after this, in 1929, the 4th Pacific Science Congress was held in Bandung at the newly opened building of the Mijnbouw or Mining Bureau. The director of the Survey, de Jongh, in the foreword to the first mapping report on Java (Koolhoven 1933), noted that this important conference was an incentive to beginning the systematic mapping of Java, not least so as to have well documented field trips for the Congress participants. It was already known that Java was richer in microfossils than Sumatra where mapping efforts had been focused, but Java also appeared to have more facies variation, and close co-ordination between field geologists and palaeontologists was considered crucial. Beginning in 1928, this work, together with similar efforts in Borneo, found many of the sections used to develop the Letter Stages. It was also this mapping work that correlated, then replaced, the field geologists “e 1 to 3” and “n 1 to 5” mapping stratigraphy with Letter Stages. The “e” and “n” annotation however continued as map symbols until the 1940’s, and modern readers of these maps appear to have forgotten the annotation is shorthand for larger foram and mollusc biostratigraphy.
The correlation is;
e1 = contains Assilina (=T.a)
e2 = No Assilina, but Pellatispira pressent (=T.b)
e3 = lower part contains Nummulites fichteli-intemedius, and an upper part has the Lepidocyclines, the top being based on the presence of Spiroclypeus s.l. (inc. Tansinhokella); T.c to basal T.e. Effectively most of the Oligocene, and it was called this by the field geologists.
n1 = Most of T.e, on Spiroclypeus and Miogypsinoides.
n2 = T.f, with Alveolinella, C. annulatus, Lepidosemicyclina, mid Early to mid Middle Miocene, both Lower and Upper T.f in the original scheme.
n3 = Lepidocyclina of the trybliolepidine type and molluscs 20-45% extant, later Middle and Late Miocene (Upper Tf of modern usage)
n4 = considered lower Pliocene with 45-60% extant molluscs, the trochid gastropod Solariella amblygoniata possibly being diagnostic.
n5 = later Pliocene and younger based on beds with either mammal fossils, or the molluscs Natica sulcifera, Turris gendingganensis, Ringicula arctatoides and Pecten tjaringinensis.
This field geologist’s scheme was devised by workers who were both geologists and capable palaeontologists such as Tobler and Rutten. The “e” and “n” notation coming from the terms “Eogenic” and “Neogenic”, where the former is equivalent to the Nummulitique of Haug in Europe

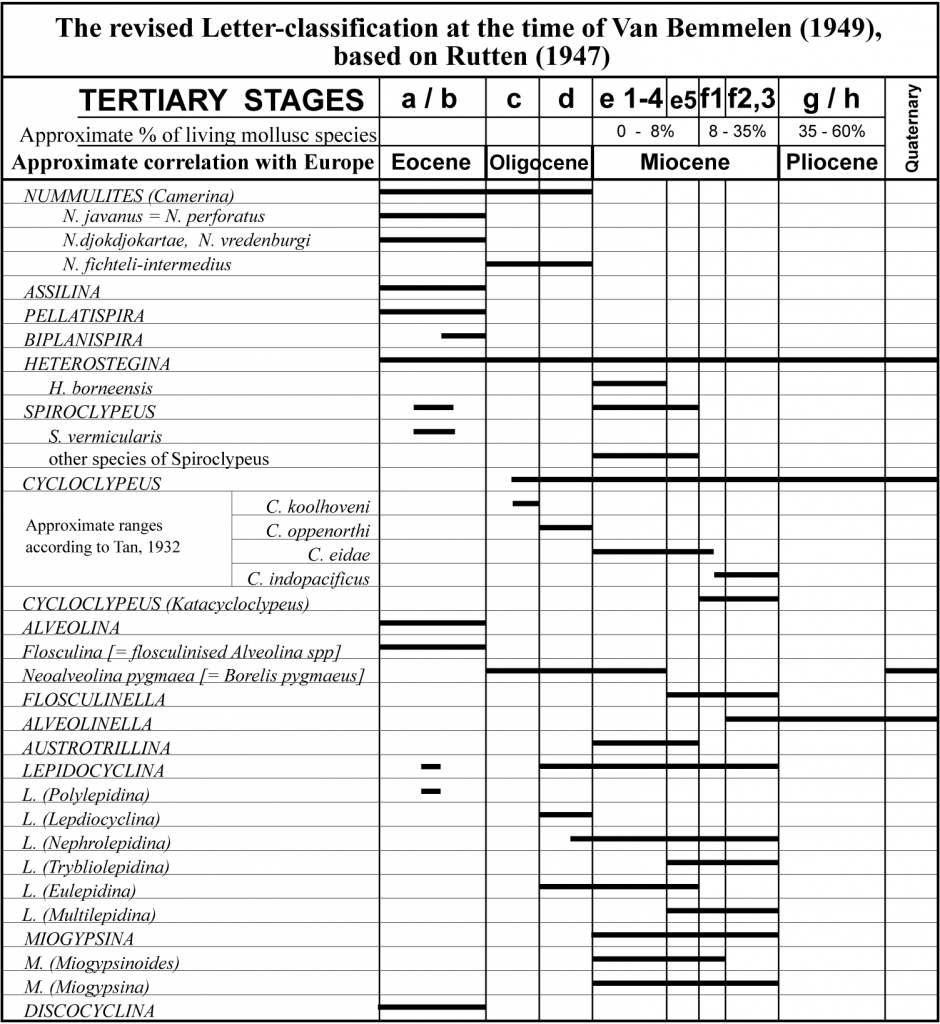
In 1931 Leupold and van der Vlerk published a detailed revision of the Letter Classification. This new version had sixteen pre-Quaternary divisions or subdivisions to replace the previous seven. The primary divisions were a through to h and for the first time were were named Stages although they are much closer in principle to the European Epochs or Series. Subdivisions, such as T.a1, T.a2 etc., were called zones. It is significant to note that the new T.g and T.h units were not defined on larger foraminifera and are not detailed in the text of this paper. The accompanying figure however noted the percentages of extant molluscs for the T.g, T.h1 and T.h2 divisions.
This 1931 paper marked the peak of the Letter Classification. Over the next twenty years the scheme was periodically modified, with a tendency to lump the 1931 “zones” rather than split them. By 1955 Van der Vlerk (cf. Rutten in Van Bemmelen, 1949; and van der Vlerk 1955) had reduced the number of pre-Quaternary divisions to eight. Tertiary a and b, as well as T.e1 to T.e4 had been reduced to single units after criticism from Tan (1939) and others, as were T.f2 and T.f3 and T.g and T.h.
This apparent failure to develop the zones to a high resolution zonation is due to the inability to consistently recogise individual species’ evolution and extinction datums in a shallow marine, hiatus-rich and facies controlled sedimentary setting.
Nethertheless for more than a decade after Leupold & van der Vlerk’s 1931 paper the larger foraminifera were the premier biostratigraphic tool in the Dutch East Indies, and were also being used more elsewhere. However in mid 1942 the Japanese Imperial forces took over the Mijnbouw and field-work virtually ceased. This was the end of the efforts of the Opsporingsdient or exploration department of the Minng Bureau, but not by the main researchers, who returned to academic posts in the Nederlands to write up their work. A great loss were the premature deaths of Tan Sin Hok in 1945 and Rutten in 1946. After this time geological work in Indonesia passed into the hands of the new Indonesian geological survey and the BPM [Bataafsche Petroleum Maatschappij] and related oil exploration companies, who did not always publish their findings.
In Irian Jaya in the 1950’s and 60’s, the scheme used by the NNGPM [Nederlandsche Nieuw Guinea Petroleum Maatschappij] was based on an internal report by Mohler which did not differ much from Rutten (1947) and van der Vlerk (1948), except to extend the range of Austrotrillina higher as the particular facies preferred by this genus is found much more often in younger rocks in east Indonesia. The quality of material in this area allowed NNGPM to re-separate T.a from T.b. The NNGPM team also started integrating early planktonic foram data with larger foram ranges, but this was hampered by the rather indurated limestone facies that dominated Irian Jaya through much of the Tertiary, plus having only the late 1950’s rudiments of the developing science of planktonic biostratigraphy.
In western Indonesia most oil was being found in the Miocene and it is this section that was most studied by the BPM. In 1957 Muhar completed an internal report that was never published, but nethertheless was an important contribution on the Oligo-Miocene section. In this work he carefully observed the mid Oligocene to Early Miocene of Java and the evolution of the Neorotalia to Miogypsina lineage, and the development of Spiroclypeus s.l. from Heterostegina borneensis. This allowed subdivision of the T.e Stage by selecting the best evolutionary events in the complex range chart of Leupold and van der Vlerk (1931).

In 1962 Eames, Banner, Blow, and Clark reviewed the basic principles of mid-Tertiary stratigraphy in a landmark contribution, making considerable advances in sorting out the complex stage and biozonal systems from Eocene through to the Miocene, and correlating most of this with a planktonic stratigraphic framework (the planktonic review only covered sediments up to the mid Early Miocene). This work included a world-wide review of mid Tertiary larger foraminifera and a consideration to the Far East Letter Stage system. Two unconformities were suggested for the Letter Stages; first between T.a1 and T.a2, with no Early Eocene represented in the Letter Stage system. Secondly an unconformity was noted in mid-Oligocene times.

The Eames et al (1962) review mentioned the Papuan Stages, which were updated by Clark and Blow (1969). These were a parallel scheme to the Letter Stages developed by field work in Papua New Guinea.
In 1970 the Letter Classification was reviewed by C.G. Adams who extended the breadth of coverage of larger foram genera and added notes on some planktonic zonal datums. In 1984 this same author detailed the Neogene with additional planktonic zonal control and clarification of taxonomic concepts.
Adams took a simple biozonal view of the Letter Stages and considered to top of Stage Tf as the highest Lepidocyclina, known to be found in rare occurrences as high as the Mio-Pliocene boundary (Adams 1984). However the original concept of Tertiary f was first defined as the interval between the end T.e extinctions [Spiroclypeus and Eulepidina] and the mass extinction of Lepidocyclina, all Miogypsinids, Flosculinella bontangensis, Cycloclypeus annulatus and Austrotrillina (van der Vlerk & Umbgrove 1927). This was correlated with mollsuc stratigraphy to what we now regard as being within the Middle Miocene. From review between 1931 (Leupold and van der Vlerk) and 1962 (Eames et al) it was noted that this mass extinction was to some dgree gradual with markers such as Austrotrillina and perhaps Flosculinella did not range as high as the more cosmopolitan Miogypsina (including Lepidosemicyclina) and the Lepidocyclina species, such as L.(N.) ferreroi which all abruptly disappeared in mid Middle Miocene times. However it was Adams (1970) who recognised that the range of Lepidocyclina extended well after the extinction of Miogypsina, as young as base Pliocene. Adams therefore extended T.f to the base of the Pliocene, irrespective of the faunal turnover in mid Middle Miocene times that was its original concept. As the Letter Stages are assemblage zones, not defined on individual evolution / extinction datums this was not an incorrect approach. However this demoted an extinction and faunal turnover event from occuring at an assemblage stage boundary to within such a stage.
THE LATEST SYNTHESIS
In the early 2000’s Tony Allan of CSIRO and Peter Lunt (Lundin Energy in Jakarta) carried out field work at some of the classic Java sites for Td, Tc to top Te, including the NE Java sites studied by Muhar (1957) and van der Verk and Postuma (1967), as well as samples archived at the Geological Survey, with planktonic foraminifera and nannofossil zonal control (with Alaa Baky) as well as strontium and larger foraminifera ages. This resulted in a manual produced for the Geological Survey short course on Micropalaeontology (Lunt and Allen, 2004). There were some revisions to this in Lunt (2013b) and van Gorsel et al (2014). The age ranges of Adams are modified only very slightly but the calibration to standard geological times scales was greatly enhanced, as were the biometrically observed evolution of Lepidocyclina (Nephrolepidina), Miogypsinoides, Miogypsina and Cycloclypeus.

The regional transgression in Tertiary A3
In Sundaland and surrounding areas mid- to later Eocene sediments usually directly overly basement or meta-sediments. There are a very large number of records of Middle Eocene or Ta3 faunas in such transgressive sediments, far more than the very rare observations of Ta1 and Ta2 faunas. In some areas the transgression took slightly longer and it may be Late Eocene (T.b) before the first marine sediments are observed.
A typical example of this succession is the detailed work of Adams (1965) on the Melinau Limestone in Sarawak, which has eroded slatey shales and quartzitic sandstones of the Mulu formation being overlain by thick limestone. The lowest thin limestone has T.a3 faunas, identified by the giant Nummulites javanus, and an absence of T.b forms. Overlying this are thick T.b (1,000-2,000 feet), T.c & d (combined up to 2,000 feet), and a similar 1,000 to 2,000 feet of T.e limestone.
On the Ta3 / Tb Letter Stage boundary
The transition from Tertiary a to Tertiary b saw the extinction of the genera Assilina / Planocamerinoides, Alveolina, and all the large complex and strongly dimorphic Nummulites. The complex Nummulitids were a rock-forming bioclasts and the rapid extinction of so many species was an important biological event.
Following the influential paper of Adams (1970) many workers have identified T.b on the presence (total range zone) of Pellatispira and the related Biplanispira. In addition to Adams, Eames (in Davies 1971) noted no known co-occurrences of Assilina with its possible descendants Pellatispira and Biplanispira. However there are several old references from Indonesia with Pellatispira occuring before the end T.a extinctions. Umbgrove (1928) is quite specific in this matter, noting several locations where Pellatispira occurs with Assilina / Planocamerinoides and T.a Nummulites such as strongly granulate species and N. bagelensis. He also notes Alveolina in a few of these samples. The figure included here (Image 10) from the T.a beds of Watu Perahu in Jiwo, C. Java, has common N. javanus in the same facies. The image shown here has several specimens of Pellatispira aff. provalei, with N. bagelensis and other granulate forms the secondary T.a marker Linderina, as well as the planktonic foraminifera Acarinina and Morozovella, demonstrating that Pellatispira not only evolved with T.a3 but that this was within the Middle Eocene (Zone P14 and older).
Elsewhere other T.a limestones with age markers such as Orbitolites are reported to contain Pellatispira, for example in south Sulawesi (Dollfus, 1915).
Until we have a detailed taxonomy of the Nummulites species from the Eocene showing the multiple extinctions, the end T.a extinction event will always be under-represented on range charts. The easiest way to emphasise the severity of the T.a3 to T.b transition is the fact that it takes only a few moments to distinguish a thin section of T.b from T.a3 age as the whole fauna has shifted in character, even though we currently lack the species level taxonomy to adequately describe this transition.
There is one location where part of the transitional T.a3 to T.b fauna is known. This is from the Cimuncang site of Koolhoven (1933) in West Java. Originally this location was the single site in West Java dated as T.a on the presence of Assilina. Recent unpublished work has relocated the single limestone bed in a mudstone sequence. In the 1930’s Koolhoven and the Mijnbou were not able to carry out planktonic biostratigaphy and even now it is hard because of the slightly mineralised and baked nature of the lithologies. However the few samples with good planktonic faunas lack Morozovella and Acarinina, and are dated as Zone P15. Elsewhere in the region (eg. the Cipatulah well to the south) this mudstone-dominated sequence has a similar age. The limestone at Cimuncang has a flat, evolute Operculina species but no sign of Assilina. It does contain fairly large, moderately dimorphic and slightly granulate Nummulites species, including N. hoogenraadi (Doornink), the type location for which is nearby in a section lacking any plantonic fossils. In both the Cimuncang and the N. hoogenraadi type location the limestones contain advanced species of Pellatispira, which have a much larger late adult or gerontic stage compared to the upper T.a3 forms illustrated from Watu Perahu at Jiwo Hills. Both the early T.b locations also have common Palaeonummulites gerthi (Doornink), a species possibly restricted to the T.b Stage. This survival of a few granulate species into T.b is not without precedent as Eames (in Morley Davies 1971) notes that in western Tethys a single granulate species (N. yawensis) survives after the extinction of most granulate species.
Tertiary B
Following the end T.b there is a radiation of new larger foraminifera types, particularly in the Pellatispirine and Heterostegine lineages. Fairly large, flat but involute Operculina species are noted in T.a 3 (Image 10) but in the T.b subdivision of chambers evolved in a number of different ways to give rise to simple Operculina eniwetokensis, with pinching-out of adult chamber growth in the median plane, the rare Grzybowskia(Christmas Island, Lunt 2003), true secondary septa in Heterostegina (Vlerkina) which developed into Tansinhokella. This radiation of the Heterostegines parallels a similar record in the western Tethyan area (Banner and Hodgkinson, 1991)
The evolution of the Pellatispines saw a rapid development of the adult or gerontic stage and the separation of the Biplanispira lineage. The work of Hottinger et al (2001) has greatly improved out knowledge of the morphology of these complex forms but the new taxonomy has not yet been applied to the sections in the Indo-Pacific. Within P15 times (from material dated with Porticulasphaera semiinvoluta from Sangiran, Gamping Barat and Nanggulan in Central Java) forms had arisen with the spiral stage reduced to a single whorl and the bulk of the test being composed of minor chamberlets or cubiculae derived from an evolutionary exaggeration of the late adult stage of ancestral forms.
Forms that were present in Ta but become more common in T.b are Amphistegina and Asterocyclina, and possibly the abundance of corals.
On the Australian Plate, which was much further south in Eocene times, the Pellatispirines are virtually unknown and T.b is often characterised by the species Lacazinella wichmani, which appears to be restricted to the Australian plate. This is known to occur in with Nummulites javanus and Alveolina (T.a), as well as with Nummulites fichteli (cf. Adams, 1970, Bursch 1947, confirming T.c) but the majority of records of this species are considered to be equivalent to the T.b Stage (Lunt 2003).
On the T.b / T.c Letter Stage boundary
At the moment it is widely assumed that the T.b-T.c boundary is equivalent to the top of the Eocene as defined by other means, most commonly planktonic foraminifera and nannofossils. The planktonic marker datum planes are correlated to be just below the base of the normal polarity of magnetostratigraphic chron C13 (ref.). However just within the base of the normal polarity of chron C13 is a marked oxygen isotope shift that has been interpreted as the result of an abrupt cooling event (Mei, 1991). It is possible that the shallow marine, tropical carbonate faunas were catastrophically affected by such an abrupt cooling .
On any stratigraphic range chart for larger forams, this T.b to T.c transition stands out as one of the most significant extinctions. Most notably the dominant Eocene larger foram, Discocyclina, and its relatives such as Asterocyclina, abruptly became extinct, as did the varied forms of Pellatispira and Biplanispira. The Eocene Heterostegina (Vlerkina) to Tansinhokella lineage seems to have become extinct, and is therefore unrelated to the Oligocene forms with the same name. This example of iterative evolution has two lines of evidence. Firstly the Early Oligocene has no records of Tansinhokella, just Planostegina and rare, slightly involute Hereostegina (cf. Tan Sin Hok 1932). Secondly a gradual evolutionary succession from these slightly involute Heterostegina forms to Tansinhokella is seen in mid-Oligocene sediments, ultimately evolving into Spiroclypeus s.s. in latest Oligocene (Lunt and Renema 2014).
Tertiary C
The next larger foram association (T.c), lasted about 3 Ma. This was the period after the terminal Eocene extinctions but before a migration of new forms arrived from Central America. It is during this period that Sr isotopic dating becomes practical and is now being applied to date these and younger limestones.
Carbonate faunas with foraminifera from this period are characterised by Nummulites fichteli-intermedius with their distinctive reticulate septal traces. Other forms found in this Stage include large species of Planostegina (P. bantamensis and praecursor Tan). Some genera are found more commonly in this stage than before or after, but are not index species. These include Borelis pygmaeus and Halkyardia.
The transition from Letter Stage Tc to Td
The original definition of van der Vlerk and Umbgrove (1927) defines the Tertiary d on the co-occurrence of Nephrolepidina and Eulepidina, together with Nummulites. The first appearance of the family Lepidocyclinidae in the Indo-Pacific region, from the much older central American stock, has been argued to be diachronous by Adams (1984) because of assumed slow migration of the genera from America and across Tethys. This model would suggest the base of T.d is a poor zonal boundary.
In recent years it has become acceptable to lump Tertiary C and D together, hence the informal name “CD limestones” for Early Oligocene carbonates in eastern Java. A more extreme opinion suggested in some oil industry reports in the late 70’s and 80’s was that the T.d stage was an artifact of reworking of Nummulites following the large mid-Oligocene eustatic sea level fall. This followed the influential papers on eustacy and stratigraphy by Vail and others at that time. Note that no such cited cases had any other genera reworked; that is, erosion from the proposed 150 metres sea level fall only seemed to affect Early Oligocene and never seemed to reach Eocene strata.
New work in the Cimanggu section of West Java (Lunt, 2013b) dates the T.c to T.d transition and also notes that a third taxon, Neorotalia mecatepecensis, may all have migrated from Central America at this time. The arrival of these forms was at about 32 Ma, near top planktonic zone P19, which appears to be before the mid-Oligocene sea-level fall. This major sea-level fall was proposed to have been eustatic by Vail et al. (1977), with revisions by Haq et al (1987) who dated it at 30 Ma, hence its popular title of “the 30 million year sea-level fall”. Later work on the Tertiary time scale culminating in Berggren et al (1995; the same time scale used here) re-dated the level given in Haq et al. at Chron C10n, intra- P21 and NP24 event to about 28.5 Ma.
There is little published data precisely dating the Vail / Haq mid-Oligocene event. Some early efforts such as Olssen et al (1980), on well cuttings from the Atlantic margin of the USA, suggested a slightly older Early Oligocene age although a revision of this work compared to the Irish Atlantic margin by Miller et al (1985) suggested the unconformity studied was closer to Vail’s mid-Oligocene level (c. 31 Ma). The problem with such studies was their reliance on cuttings-samples, as only deep boreholes penetrate the para-conformable expression of the event, and also the weakness of planktonic biostratigraphic zones in the early to mid-Oligocene.
More recently Kominz and Pekar (2001) studying the profile of the eastern seaboard of the US Atlantic margin used strontium dating and backstripping techniques to study Oligocene sea levels. Using the same time scale as Beggren et al (1995) these workers did not recognise a single outstanding eustatic event in mid-Oligocene times. Instead their data (cf. their figures 2 and 8) show a moderate sea level fall of about 45 metres between 27.9 and 28.3 Ma (both ± 0.7 Ma), and only smaller, fluctuating rising and falling, sea-level changes in the period 31.5 to 33 Ma.
This data suggests that the outstanding major (c. 150 metre) sea-level fall shown on oil-industry sequence stratigraphic charts in mid-Oligocene times, if it ever existed, was not likely to have been the cause of the mass-migration of central American larger foraminifera into the Mediterranean and Indo-Pacific areas. Also the migration reached the Far-east at an older date than previously estimated, so Adam’s (1984) hypothesis of slow, diachronous migration over millions of years is probably not correct.
The top of Td
Unlike the sudden appearance of the Lepidocyclinids at the base of T.d, Nummulites seems to disappear more gradually. There are records of this species as high as the extinction of Chilguembelina cubensis in field samples from the center of the Kujung Anticline in NE Java (28.5 mybp), and as young as 28.3 Ma at the Pelang limestone C-E Java (Pelang Lst 87Sr/86Sr = 0.70804, NBS987 = 0.710235, Age of 28.3 mybp, combined precision & correlation error 27.59 to 28.89 Ma. Center of Kujung Anticline T.d / top P21(a) sample: 2K1/9/21, 87Sr/86Sr = 0.70800, NBS987 = 0.710235, Age of 29.3 mybp, combined precision & correlation error 28.58 to 29.95 Ma.). However in the Kujung anticline we have found single specimens of Nummulites slightly higher into the basal Late Oligocene, but not yet after the evolutionary appearance of Tansinhokella (base Te2-3). However Hashimoto et al. (1977) working on the Bugton Limestone of east Mindoro in the Philippines have clearly illustrated Nummulites next to Tansinhokella. This unique sample was a loose boulder of limestone, so while their photograph is clear and shows no difference in preservational type, the location or strata cannot be resampled.
Therefore it is likely that the disappearance of this very important genus was gradual and took place over one or two million years. If Nummulites did become extinct after the evolution of Tansinhokella then Letter Stage Te1 is not strictly viable, although the most significant reduction in Nummulites abundance at about 28.5 Ma remains a useful biostratigraphic datum.
Subdivision of Te
The unpublished work of Muhar (1957) of the BPM, discussed near the start of this blog, remains the best method to subdivide the T.e Letter Stage. This scheme simply traces the evolutionary development of Neorotalia mecatepecensis to Miogypsinoides and to Miogypsina, and Heterostegina (Vlerkina) to Tansinhokella. These events are approximately fixed to biostratigraphy and some Sr isotopic ages. Only the final transition from Tansinhokella to Spiroclypeus remains uncalibrated.
Many workers use the extinction datum of Heterostegina borneensis as a secondary index for the Te4 to Te5 boundary. In many cases this seems a reasonable approximation, however in northern Borneo (Sabah) van der Vlerk (1951, p.145) has noted H. borneensis in association with Miogypsina. This observation was also noted in the same area, in samples from the Gomantong Limestone, by Adams (1964, p.162) who also stated “It [H.borneensis] is, nevertheless, most abundant in Te1-4 strata and its occurrence with large numbers of eulepidines and very few nephrolepidines may be taken as indicating Te1-4 beds in Sabah.”
On the Upper T.e / T.f Letter Stage boundary
The position of the top of Letter Stage T.e, with respect to planktonic zones or absolute ages, is now adequately fixed, although some references are still confused. Most publications (Chaproniere 1975 & 81; Adams 1984; Jenkins et al., 1985) suggest the top occurrences of Spiroclypeus and Eulepidina are at about the N5 to N6 zonal boundary, which current estimates place at about 19-20 Ma. This is close to a major regional, tectono-stratigraphic event that dominates the stratigraphic record, with strong facies changes.

This regional geological event is characterised by a change from widespread carbonate development (a prolonged flooding period) to the appearance of a new and highly active clastic sedimentary source. In Sundaland this severe facies change includes at least a hiatus, or deposition of a substantial clastic section before the occurrence of the next carbonate related facies. Isolated reefs seen in oil exploration may have T.e carbonate separated by only a small hiatus from basal T.f carbonate (e.g. the Kujung followed by Rancak limestones in offshore NE Java) however these are coral dominated reefs, often pinacle reefs with limited foraminiferal grainstones, and they have only been encountered in oil wells with cuttings and limited core samples. They are not known in outcrop onshore. In 2021 Lunt published a review of the Rebab-1 well in Luconia which has continuous carbonate through this part of the Miocene as well as rich larger foraminifera in four cores, with supporting strontium dating. This well confirmed the empirical data from service company schemes and Adams and Chaproniere, that the top of Te was close to 20 Ma, about half a million years before the base of Zone N6.
In Irian Jaya and Papua New Guinea there is less tectonic or facies contrast during this mid-Early Miocene period but the same faunal turnover can be seen. This is known as the change from Kereruan to Kikorian faunas in Papua New Guinea. Not only do two locally important components of the larger foram assemblages disappear but there is subsequently a radiation of new forms. Note also that in eastern Indonesian and Papuan areas the relatively rare genus Austrotrillina, previously thought to be a T.e marker, is seen to continue into younger sediments. In addition some of the forms that radiated and became important in the T.f have occasional records alongside upper T.e markers (e.g. Flosculinella, and Marginopora). The distinction between T.e and T.f Letter Stages appears to becomes slightly less abrupt in areas away from Sundaland where there is maximum expression of the mid-Early Miocene tectono-stratigraphic event.
The T.e to T.f boundary is not comparable to the end T.a or T.b mass extinction events. The surviving T.f fauna is much more a continuation of the end T.e fauna rather than the major and widespread T.a to T.b, or T.b to T.c faunal turnovers
Tertiary F
The Tertiary F Letter Stage has, by far, the most confusion in its useage between generations of workers. The top of the Stage has shifted considerably, and the two or three subdivisions also have inconsistent definitions.Therefore Tf.3 of Adams (1984) is T.g of workers in the 1930’s and 40’s; and the old Dutch Tf, 1,2,3 are very different to Adams’ divisons of the same name.
Tertiary f was first defined as the interval between the end T.e extinctions [Spiroclypeus and Eulepidina] and the mass extinction of Lepidocyclina, all Miogypsinids, Flosculinella bontangensis and Austrotrillina (van der Vlerk & Umbgrove 1927). By 1931 Leupold and van der Vlerk considered that Austrotrillina became extinct slightly before the others [i.e. within Tf]. In 1962 Eames et al. noted that Flosculinella also did not range as high as the more cosmopolitan Miogypsina and Lepidocyclina. However it was Adams (1970) who recognised that the range of Lepidocyclina extended well after the extinction of Miogypsina. By 1979 Adams et al. had found Lepidocyclina as young as base Pliocene. Adams therefore extended T.f to the base of the Pliocene and first established Lower T.f [up to the extinction of Miogypsina and a few rarer species] and Upper Tf to the extinction of Lepdiocyclina. As the Letter Stages are assemblage zones, not defined on individual evolution / extinction datums this was not an incorrect approach. However this demoted a mass extinction and faunal turnover event from occuring at an assemblage stage boundary to within such a stage. It also deviated from the original Dutch concept of the Letter Stage, which was based on this mass extinction.
We can be sure that this was the intention of the early workers as their foraminiferal stratigraphy was integrated with molluscan stratigraphy, often sampling the same formations. The Top T.f of the early workers was the division between molluscan faunas with less than 35% extant species, and those with more. Examples of the older Molluscan Stage, called the Rembangian and Preangerian by Oostingh (1938, also in van Bemmelen 1949) contain Miogypsina, Cycloclypeus annulatus etc. and the type locations for both these stages are now dated with modern planktonic biostratigraphy as latest Early and base Middle Miocene respectively (pers. obs., unpublished). The molluscan stage with 35-50% extant species (Tjiodeng or Ciodeng Stage) has 42% extant species at type locality in W Java, which was considered Late Miocene by Martin 1919). The Genteng Beds of SW Java are also in Oostingh’s Ciodeng Stage, and these beds overly the Bojongmanik Fm. which contains limestone of T.f3 age (trace Lepidocyclina with Alveolinella) which has recently been dated as between 10-11 Ma by strontium isotopic methods. Another T.g location are the Lawak Beds in Central Java, such as the lower NN11 sample at Margamukti in north central Java (Lunt 2013b).
As the works of Adams (1970 and 1984) are so widely quoted it would be confusing to revert now to the original concept of Tf. However it should be stressed that the original Tf to Tg boundary was first defined as a mass extinction in mid-Middle Miocene times.
This important biostratigraphic event is also an important faunal turnover for other carbonate facies organisms. The result was that shallow marine biohermal limestones changed from mixed coral and coralline algal boundstones with larger foram grainstones to dominant coral reefs with a marked increase in Halimeda green algae and molluscs. The algae often preserved as recognisable plate fragments, or as an increase in micrite and fine bioclastic products from the early breakdown of its aragonitic needles. Larger foraminiferal grainstones are generally rarer after this event, most often as as deeper photic Cycloclypeus facies, or minor Operculina / Amphistegina or Alveolinella calcarentites.
References
Adams, C. G., 1970. A reconsideration of the East Indian letter classification of the Tertiary. Bulletin of the British Museum of Natural History (Geology) 19, 1-137
Adams, C. G., 1984. Neogene larger foraminifera, evolutionary and geological events in the context of datum planes. In N. Ikebe & R. Tsuchi (Eds.), Pacific Neogene Datum Planes, 47-68. Tokyo: University of Tokyo Press.
Chaproniere, G. C. H., 1975. Palaeoecology of Oligo-Miocene larger foraminiferida, Australia. Alcheringa 1, 37-58
Chaproniere, G. C. H., 1981. Australasian mid-Tertiary larger foraminiferal associations and their bearing on the East Indian Letter classification. BMR Journal of Australian Geology & Geophysics 6, 145-151
Eames, F. E., Banner, F. T., Blow, W. H., & Clarke, W. J., 1962. Fundamentals of mid-Tertiary correlation. Cambridge: Cambridge University Press.
Jenkins, D. G., Bowen, D. Q., Adams, C. G., Shackelton, N. J., & Brassell, S. C., 1985. The Neogene: part 1. In The Chronology of the Geological Record (Memoir 10), 199-210. London: Geological Society of London.
Lunt, P., 2013a. Foraminiferal micropalaeontology in SE Asia. In A. J. Bowden, F. J. Gregory, & A. S. Henderson (Eds.), Landmarks in Foraminiferal Micropalaeontology: History and Development. London: The Geological Society.
Lunt, P., 2013b. The sedimentary geology of Java. Jakarta: Indonesian Petroleum Association. 346 pp.
Lunt, P., 2021. Tectono-stratigraphic framework of Luconia carbonates. SEPM Special publication, Cenozoic Isolated Carbonate Platforms – Focus Southeast Asia,
Lunt, P., & Allan, T., 2004. A history and application of larger foraminifera in Indonesian biostratigraphy, calibrated to isotopic dating. GRDC Workshop on Micropalaeontology, 112
Lunt, P., & Renema, W., 2014. On the Heterostegina– Tansinhokella– Spiroclypeus lineages in SE Asia. Berita Sedimentologi 30, 6-31
Muhar, A., 1957. Micropalaeontologishe onderzoek van monsters afkomstig van het geologishe onderzoek Tuban. B.P.M. unpublished report Gr.Sb. 1770, 22
van der Vlerk, I. M., 1951. Appendix 2. in Geology of the Colony of North Borneo by Renhard, M. and E. Wenk. In Brit. Borneo Geol. Survey Bull. 1, 160.
van der Vlerk, I. M., 1955. Correlation of the Tertiary of the Far East and Europe. Micropaleontology 1(1), 72-75
van der Vlerk, I. M., & Umbgrove, J. H. F., 1927. Tertiaire gidsforaminiferen uit Nederlandsch Oost-Indië. Wetenschappelijke Mededelingen Dienst van den Mijnbouw in Nederlandsch-Indië 6, 1-31
[…] following workers over some 70 years, a single outstanding unconformity was noted between beds with Letter Stages Te4 (or slightly wider Te2-4 age in some places) and Te5 age sediments. Much of the paper deals with […]
[…] but it is usually assumed to be the Rajang Unconformity near the top of the Middle Eocene (near top Letter Stage Ta). Another assumption is that the thick fuvio-deltaic siliciclastics see in many offshore wells […]
[…] On Cycle I, Sarawak – The Orbulina datum on The Letter Stages of SE Asia […]