See also the post on environment of deposition from benthic foraminifera
Determining environment of deposition is important for tracing facies, and natural successions following Walther’s Law; and also the inverse – of recognising sudden breaks in a normal Waltherian succession. A non-Waltherian contact is an unconformity defined by a distinct shift in sedimentary systems rather than bedding geometry at a horizon, which will changes with position on the same contact.
Many properties of facies are related to water depth, which has roughly linear relationships to energy, light and several other properties that affect sedimentary processes and fossil assemblages.
Planktonic foraminifera; under-used in environmental analysis
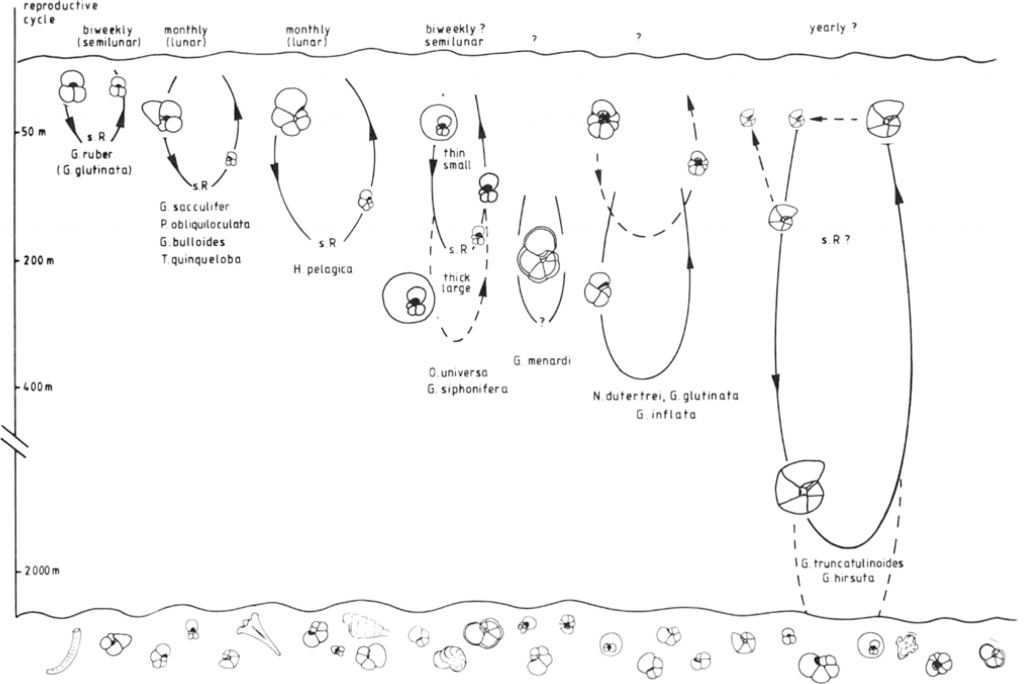
There is both biological and empirical evidence linking abundance and diversity of planktonic foraminifera to water depth. While planktonic foraminifera live most commonly in the upper 50 m of the oceanic water column (Hemleben et al., 1989; Gupta, 2002), they require greater water depths to complete their life cycle and reproduce. For some species this may include migration below the thermocline, as deep as several hundred metres. Consequently to have both high planktonic abundances in sediments, and a death assemblage with high species diversity, requires deep marine conditions. Hemleben et al. (1989, chapter 10.4) and Zheng and Fu (1994) summarise modern instances where planktonic foraminifera only become common in sediments many tens to a few hundreds kilometres away from land, and in water depths greater than 200 m.
Sea floor sampling in SE Asia (Keij, 1964; Biswas, 1976; Boichard et al, 1985; Zheng and Fu, 1994; Isnaniawardhani et al., 2003; Szarek et al, 2006, 2009, Wang et al. 2014), as well as compilations from well samples by van Gorsel (1988), Kadar et al. (1996) van Gorsel et al. (2014) support, without exception, the linear relationship between planktonic foraminifera abundance and water depth, using the metric of a planktonic to benthic foraminiferal ratio (P:B). In the EPGL facies this P:B ratio is typically 80-90%.
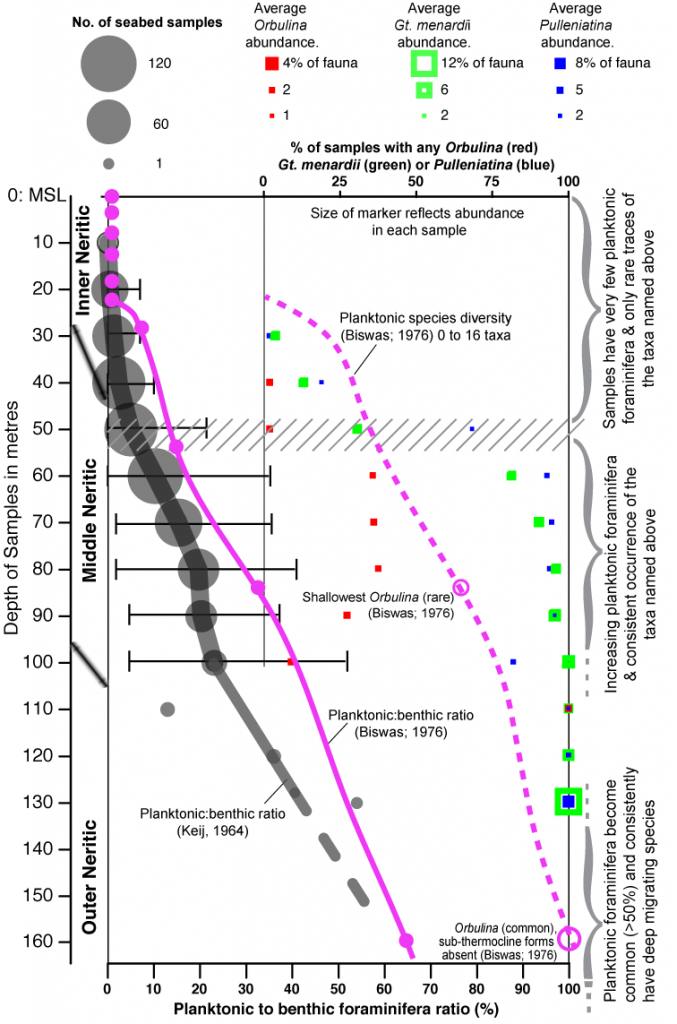
The summary figure above is based on the tabulated data of Keij (1964), with the trends of Biswas (1976) added. In inner neritic settings (to about 25 or 30 m water depth) the P:B ratio is 0 to 10% and these are of simple Globigerina and Globigerinoides forms, often juveniles. In middle neritic settings (extending down to about 100 m water depth) the ratio gradually increases to as much as 40%. A rule of thumb derived from this empirical data is that the P:B ratio only exceeds 50% in samples deposited in approximately 150 m water depth or more. Only the data from Szarek et al (2006, 2009) deviates slightly from this, with samples they note to be unusually rich in planktonic forms and a P:B threshold of 50% at about 100 m water depth, although this is in a setting over 200 km from the nearest significant land masses. Taxa associated with deeper marine life cycles can be found in small numbers in middle neritic settings, but inconsistently present. This is why in shallower marine samples the nannofossils replace planktonic foraminifera as the best bio-zonal indicators. The taxa Orbulina, the menardi-form and tumid Globorotalias, and the genera Paragloborotalia, Pulleniatina and Dentoglobigerina are only occasionally found shallower than 50 m, and are common only in samples approaching 200 m water depth. Other taxa such as Sphaeroidinella, Globigerina venezuelana and the hirsuta-lineage of globorotalids are only frequent in samples from bathyal settings (>200 m; “open ocean sub-thermocline”; Aze et al., 2011).
Analysis of oxygen isotopes, which are incorporated into the test at ratios linked to water temperature and thereby depth, has confirmed this modern distribution, based on sea-floor samples and plankton netting. This correlation allows environmental preferences for extinct species to be estimated (e.g. Pearson and Wade, 2009; Aze et al., 2011, and many others). The extinct Sphaeroidinellopsis, Catapsydrax, Fohsella, Globorotalia margaritae and Globigerina binaiensis-sellii lineage, and most Hantkenina species appear to require deep outer shelf or bathyal conditions (c. 150 m or more) to be frequent or consistently present in samples. The species G. margaritae is a member of the Hirsuta lineage (ex Hirsutella of Bandy 1972; name not available), and is the marker for Zone 19/21c. This form is found in multiple wells from the EPGL, from onshore East Java (Ngasin, Gondang and Grigis Barat) to Wortel, Oyong and Maleo in the east.
The P:B ratio is an imprecise index, but it is highly repeatable in many data sets (it has accuracy) and it is considered to have high trueness when compared to independent palaeobathymetric controls (e.g. depth converted seismic maps), and there are no known exceptions where sediments with abundant planktonic foraminifera and open oceanic markers are found shallower than about 120 to 150 m (the reports of Szarek et al. mentioned above is just to common abundance, P:B 50%, at 100 m and not high diversity). In addition the occurrence of several larger foraminifera taxa in water depths less than 153m, and especially less than 50 m (Leutenegger, 1984) rules out the EPGL from being a middle shelf or shallower facies, and the diversity of planktonic forms including deep migrating, sub-thermocline, species indicates bathyal condition (>200m) were very likely.
References
Aze, T., Ezard, T.H.G., Purvis, A., Coxall, H.K., Stewart, D.R.M., Wade, B.S., Pearson, P.N., 2011. A phylogeny of Cenozoic macroperforate planktonic foraminifera from fossil data. Biological Reviews 86(4), 900-927
Biswas, B., 1976. Bathymetry of Holocene foraminifera and Quaternary sea-level changes on the Sunda Shelf. Journal of Foraminiferal Research 6, 107-133
Boichard, R., Burollet, P.-F., Lambert, B., Villain, J.-M., 1986. La Platform carbonateé du Pater Noster, Est de Kalimantan (Indonesia). Paris: Total, Companie Française des Pétroles.
van Gorsel, J.T., 1988. Biostratigraphy in Indonesia: methods, pitfalls and new directions. Proceedings from Proceedings Indonesian Petroleum Association Convention: 17, 275
van Gorsel, J.T., Lunt, P., Morley, R.J., 2014. Introduction to Cenozoic biostratigraphy of Indonesia- SE Asia. Berita Sedimentologi 29, 4-40
Gupta, B.K.S., 2002. Modern Foraminifera, 2nd edition. Dordrecht: Kluwer Academic Publishers.
Hemleben, C., Spindler, M., Anderson, O.R., 1989. Modern planktonic foraminifera. Springer-Verlag.
Isnaniawardhani, V., Suparka, E., Kapid, R., Latief, H., 2003. Nannoplankton and foraminifera assemblages and their relations to bathymetry in Madura waters. Proceedings from Eighth International Congress on Pacific Neogene Stratigraphy.
Kadar, A.P., Paterson, D.W., Hudianto, 1996. Successful techniques and pitfalls in utilizing biostratigraphic data in structurally complex terrain: VICO Indonesia’s Kutei Basin experience. Proceedings Indonesian Petroleum Association Convention 25, 313-331
Keij, A.J., 1964. The relative abundance of recent planktonic foraminifera in seabed samples collected offshore Brunei and Sabah. Brit. Borneo Geol. Survey Ann. Rept. for 1963, 146-153
Pearson, P.N., Wade, B.S., 2009. Taxonomy and stable isotope paleoecology of well-preserved planktonic foraminifera from the uppermost Oligocene of Trinidad. Journal of Foraminiferal Research 39(3), 191-217
Szarek, R., Kuhnt, W., Kawamura, H., Kitazato, H., 2006. Distribution of recent benthic foraminifera on the Sunda Shelf (South China Sea). Marine Micropaleontology 61, 171-195
Szarek, R., Kuhnt, W., Kawamura, H., Nishi, H., 2009. Distribution of recent benthic foraminifera along continental slope of the Sunda Shelf (South China Sea). Marine Micropaleontology 71, 41-59
Zheng, Z.Z., Fu, Z.X., 1994. Foraminiferal faunal trends in China Seas. In D. Zhou (Ed.), Oceanology of China Seas, 225-274. Dordrecht: Kluwer Academic Publishers.
Be First to Comment